In this section the physiological and aerodynamic behavior of the voice source will be discussed. It will be shown how the voice source is used to achieve various linguistic and paralinguistic goals.
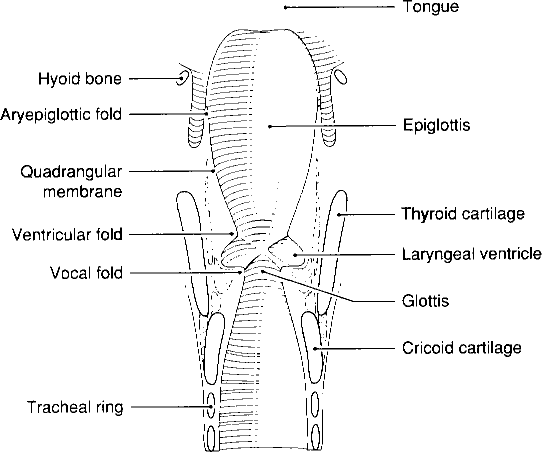
An airstream formed in the lungs passes into the trachea and finally into the larynx, which is situated on top of the trachea. The larynx is the principal structure for producing vibrations of the air stream, while the oscillating vocal folds are responsible for activating vibrations of the air . The rapid closing and opening of the vibrating vocal folds periodically interrupts the air stream and produces a glottal tone later modified in the cavities of the vocal tract. The larynx is a musculo-cartilaginous structure supported by muscles from the hyoid bone which enable it to move up and down (Fig.2). When swallowing, the larynx is moved upward and the epiglottis folds down over it, covering the entrance to the trachea in order to prevent food from entering into the lungs. During yawning the larynx is lowered and moves down in order to widen the airway.
| Figure 2. Coronal section through the larynx showing the laryngeal airway (from: Titze, 1994:16). |
The non-biological function of the larynx - the production of sound - depends on the activity of the muscles and the position of the cartilages of the larynx.
The cartilaginous framework of the larynx consists of the thyroid, cricoid and arytenoid cartilages. The cartilages and the muscles associated with the larynx are shown in Fig. 3.
The vocal folds are attached at the front of the larynx to the thyroid cartilage and at the back to the arytenoid cartilages. The arytenoid cartilages can be made to rotate in a swivelling movement and to slide apart along the cricoid cartilage on which they are located. The movement of the arytenoid opens a three-dimensional triangular space (tetrahedron) between the inner edges of the vocal folds, i.e the glottis. The movement of the vocal folds in the process of separation is called abduction. If the folds approximate, they are adducted. The movement of the arytenoid regulates the abduction and tension of the vocal folds.
The vocal folds (Fig.2 ) are located in the narrowest portion of the airway, just below the laryngeal ventricle and the ventricular folds2. The vocal folds have a layered structure. The outermost layer is a thin skin made up of stratified squamous epithelium which is 0.05 to 0.1 mm thick. Under the epithelium resides the lamina propria, usually divided into three layers: superficial, intermediate and deep. The first layer, 0.05 mm thick, consists of elastin fibres surrounded by internal fluids. Elastin fibres allow for considerable elongation. The intermediate layer is also made of elastin fibres, but they are more uniform longitudinally. The third layer is made up of collagen fibres, which in opposition to elastin fibres, are not extensible. The intermediate and deep layers together are about 1 to 2 mm thick (Titze, 1994:15-17; Hirano et al., 1981). Those layers lie on top of the thyroarytenoid muscle (and muscle vocalis3), which is a major part of the vocal folds, being approximately 7 to 8 mm thick. The muscle fibres run longitudinally along the folds, but are are shaped similar to a braid.
The different properties of the layers lead to a different labelling of the structure of the vocal folds in which the fold consists of a body (a deep layer of lamina propria and muscle) and a cover (epithelium plus superficial and intermediate layers of the lamina propria). The body-cover model of the vocal folds' structure is used for the modelling of vocal fold vibration (see section 27.3). The vocal folds are approximately 11 to 21 mm long. The structure of the folds allows changes in shape and thickness, but also elasticity and/or stiffness.
| Figure 3. Schematic view of the muscles connecting the thyroid and cricoid cartilages (from: Laver, 1994: 185). |
The vibratory movement of the vocal folds can be approximated by the classic myoelastic aerodynamic theory of vocal fold vibration (van den Berg, 1958). According to the Bernoulli law:
If the volume of airflow is constant, the velocity of flow must increase at the region of the constriction, but with a corresponding decrease of pressure at the constriction. This means that the vocal folds are sucked together by negative Bernoulli pressure in the glottis. This action is only possible if the glottis is sufficiently narrow, if the airflow is sufficiently high and if the medial surface of the vocal fold is soft enough to yield (Titze, 1994:81). The stricture between the vocal folds (stricture of the glottis) is then followed by a build-up of subglottal pressure during closure, causing the folds to move outward and the glottis to open (ibid.). The movement continues until the elastic forces of the tissue retard the motion and reverse it. The folds move back together again and the next cycle of movement begins.
The theory does not explain all aspects of vocal folds oscillation (see Titze, 1994:82 for discussion) and more elaborated models have been proposed. Ishizaka and Flanagan (1972) proposed a two-mass model of vocal fold vibration. Titze (1988) proposed a body-cover model of oscillation with a surface wave propagating in the cover. In both models (which will be discussed in detail in chapter V) the driving force for oscillation comes from a non-uniform movement of the top and bottom edges of the vocal folds which move with a small delay (Titze, 1988). The build-up and collapse of the airflow in the glottis and the vocal tract are then delayed with respect to the tissue movements. This causes a non-uniform pressure distribution at the glottis during the vibration cycle.
During the movement4 of the articulators the coupling between voice source (glottis) and resonator (vocal tract) varies. From the body-cover model it follows that even in the case of a varied coupling between voice source and resonator the larynx is still able to control phonation independently from articulation. This fully supports the linear source-filter model of speech production (Fant, 1960).
The position of the arytenoid cartilages plays an important role in all modes of phonation. The cartilages are moved by several muscles. The muscles of the larynx are often divided into two groups:
· intrinsic muscles interconnecting the cartilages of the larynx,
· the extrinsic muscles connecting the larynx to the surrounding structures.
The following muscles are active during phonation 5:
1. The cricothyroid muscle, which runs from the cricoid to the thyroid cartilage and elevates the cricoid and lowers the thyroid, thereby shortening the cricothyroid space and lengthening the vocal folds. This lengthening causes a change in the stiffness of the vocal folds, which in turn changes the duration of a period of vocal fold vibration.
2. The posterior cricoarytenoid muscles, which start on the posterior surface of the cricoid cartilage and run upward and laterally to the arytenoid cartilage. They rock the arytenoid, pulling the vocal folds apart, causing their abduction (Koreman, 1996:5).
3. The lateral cricoarytenoid muscles originate at the muscle of the arytenoid and run to the sides of the cricoid cartilage. While contracting, they rock the arytenoids, causing the mucous part of vocal folds to approximate.
4. The interarytenoid muscles run horizontally between the two arytenoids and adduct the vocal folds. This supports the lateral cricoarytenoid muscle in closing the posterior part of the glottis.
The intrinsic muscles of the larynx are presented in Fig. 4.
| Figure 4. The intrinsic muscles of the larynx
as shown in (A) lateral, (B) posterior,
and (C) superior views (from: Orlikoff & Kahane, 1996) |
2.the ventricular folds are also called false vocal folds.
3.the muscle vocalis is often regarded as part of the thyroarythenoid muscle.
4. the forces acting on the vocal folds depend on the shape of the vocal tract, thus a changed place of articulation may cause modification of the the mods of the folds' vibration. See section 24 for details.
5. for detailed description the reader is referred to Zemlin (1988) and Pesàk (1990).
![]()
![]()
![]()
![]()